脂肪酶的结构和作用机理
化学先生 / 2019-08-02
脂肪酶本身就是一个手性分子,近十年来,随着生物化学和分子生物学技术的发展,对其活性中心结构、作用机理等的了解日益深人,取得了令人瞩目的进展。脂肪酶立体选择性分子基础的系统阐明,对脂肪酶在不对称合成中的应用和基础理论研究是十分必要的。
1:脂肪酶和酯酶的催化活性位点
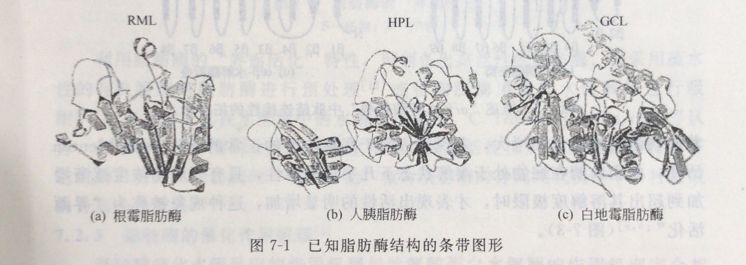
到目前为止,已经采用X射线衍射的方法对11种不同来源的脂肪酶和酯酶的结构进行了研究,发现尽管它们的氨基酸序列和分子大小相差很大,但所有的脂肪酶和酯酶都以相同的方式进行折叠并具有相同的催化位点。通过对各种脂肪酶和酯酶的结构相似性进行归纳,这种特有的折叠方式被命名为“a/β-水解酶折叠”(图7-1)。
该结构包含有一个被a~螺旋包围着的,以平行β片层结构为主的核。肽链通过这种折叠方式产生了一个丝氨酸组氨酸天门冬氨酸(或谷氨酸)三元催化位点,亲核的丝氨酸位于平行β-片层和a螺旋之间的发夹结构上,组氨酸和天门冬氨酸(或谷氨酸)位于丝氨酸的同一面,几个氨基酸残基位于丝氨酸的另一面,形成一个稳定的氧阴离子孔洞 (图7-2)。
不同来源的脂肪酶的结构差异主要在于肽链形成的折叠环的数目和位置不同,如RML这样的小分子脂肪酶,只由一个a螺旋形成覆盖活性位点的盖结构;而CRL这样的大分子脂肪酶,却由两个a-螺旋形成覆盖活性位点的盖结构。
2:脂肪酶的界面活化
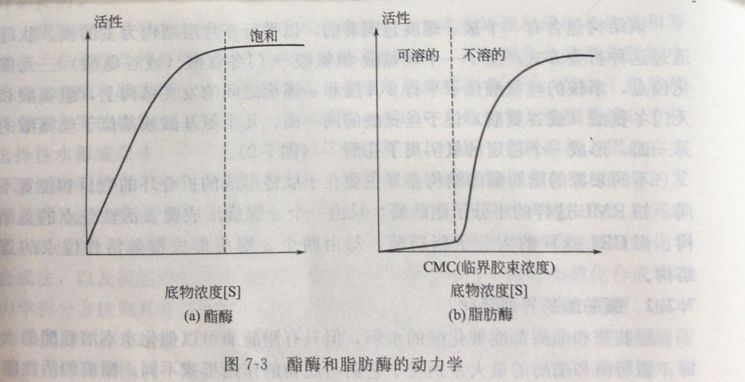
脂肪酶和酯酶都能催化酯的水解,但只有脂肪酶可以催化水不溶性酯的水解。脂肪酶和酯酶的最大差别在于它们与底物的作用形式不同。酯酶的活性随着底物浓度的增加而增大,表现为依赖于底物浓度的正常的MichaelisMenten活性,而脂肪酶在底物处于溶解状态下几乎没有活性,只有当底物浓度逐渐增加到超出其溶解度极限时,才表现出活性的明显增加,这种现象被称为“界面活化”(图7-3)。
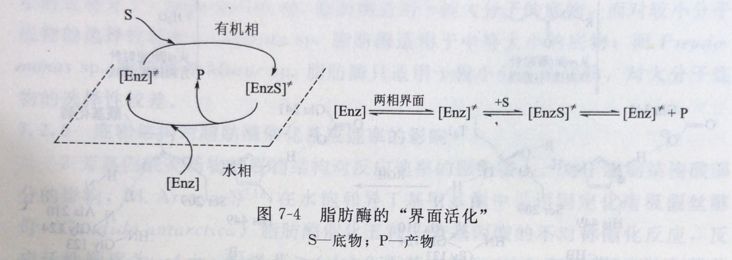
“界面活化”现象的分子基础被认为是酶分子的构象变化所至,当缺乏油/水界面时,酶分子的活性位点被一个称之为“盖”的ar螺旋结构覆盖,稳定氧阴离子的几个氨基酸残基的排列位置不正确,脂肪酶处于非活性构象[Enz];当脂肪酶与两相体系的油/水界面接触时,覆盖活性位点的ar螺旋结构打开,暴露出活性位点,同时这种“盖”的打开,也使稳定氧阴离子的几个氨基酸残基位于正确的位置,脂肪酶处于活性构象[Enz]≠(图7-4)。
利用脂肪酶的“界面活化”特性,可制备出高活性的脂肪酶,如采用疏水性的有机溶剂对脂肪酶进行预处理或将脂肪酶用强疏水性载体进行吸附,可以增大其在水溶液中的水解反应活性。C. Torres与A. Bastida等认为,这同样是由于酶的分子构象发生变化,覆盖活性位点的ar螺旋结构打开造成的。另外,脂肪酶的“界面活化”也为脂肪酶的分离纯化提供了一种有效的分离纯化方法。
3:脂肪酶的催化作用机理
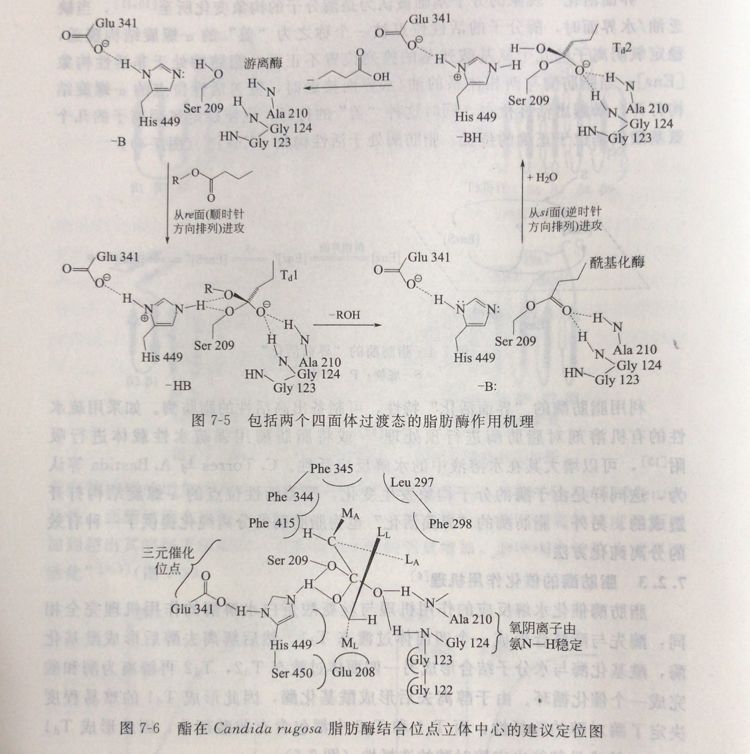
脂肪酶催化水解反应的作用机理与丝氨酸蛋白水解酶的作用机理完全相同:酶先与酯结合形成一个四面体过渡态Tal,然后解离去醇后形成酰基化酶,酰基化酶与水分子结合形成另四面体过渡态 Ta2,Tu2再解离为酶和酸完成一个催化循环。由于醇离去后形成酰基化酶,因此形成Ta1的难易程度决定了酶对醇的选择性:而Tal和Ta2中都包含有羧酸部分,因此形成Ta1和Ta2的难易都能决定酶对酸的选择性(图7-5)。
4:脂肪酶的选择性及其分子基础
脂肪酶的分子结构研究表明,脂肪酶上的酯结合位点可以划分为三个区城:位于β片层上方的ML区;疏水腔;位于a-螺旋上方的隧道区。酯在催化位点上的排列方式为:醇部分结合于ML.区和疏水腔,酸部分结合于疏水腔和隧道区(图7-6)。
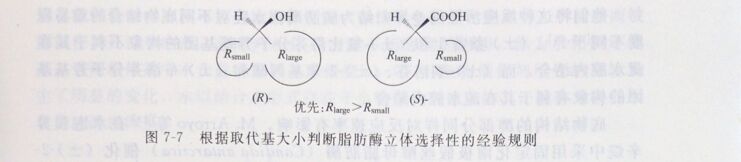
醇部分ar碳上的中等大小取代基位于ML区,a-碳上的较大取代基位于疏水腔;对于酸结合部分,立体中心被认为是位于隧道口处,a-碳上的较大取代基位于隧道区,a-碳上的中等大小取代基位于疏水腔。羧酸部分简单的结合模型如图7-7所示。
由于不同脂肪酶的酸结合位点有差别,由此可以说明为什么不同脂肪酶对酸具有不同甚至相反的立体选择性。不同的脂肪酶隧道区的长度差别很大,对于CRL,隧道区的长度可以容纳至少18个碳原子的碳链,而RML和HLL的活性中心到酶分子表面的隧道区长度却很短。因此不同的脂肪酶适用于不同大小的底物分子,Aspergillus sp.脂肪酶适用于较大分子的底物,而对较小分子底物的选择性较差; Candida sp.脂肪酶适用于中等大小的底物;而Pseudo-monas sp.脂肪酶和Mucor sp.脂肪酶只适用于较小分子的底物,对大分子底物的选择性较差。
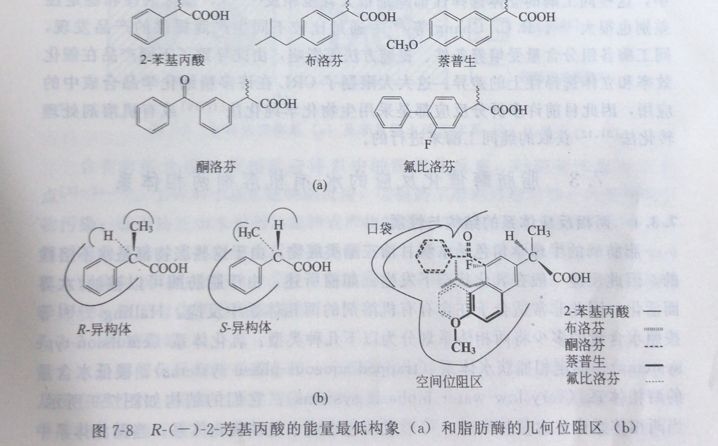
5:底物结构对脂肪酶催化反应速率的影响水西
2-芳基丙酸类药物的底物结构对反应速率的影响很大。对于底物结构酸部分的影响,M.Arroyo等在水饱和异丁基甲基酮中采用固定化南极假丝酵母(Candida antarctica)脂肪酶催化五种2-芳基丙酸的不对称酯化反应,反应活性顺序为: (士)-酮洛芬>(土)-2-苯基丙酸>(士)-布洛芬>(士)-蔡普生=(士)-氟比洛芬。Sih 等采用游离的柱状假丝酵母(Candida cylin-dracea)脂肪酶催化上述五种2-芳基丙酸甲酯和2-芳基丙酸乙酯的不对称水解反应,也得到了同样的反应活性顺序(图7-8)。
他们将这种反应话性的差归结为脂防附成水胶对不同底物结合的难易程度不同。(士)-蔡普生和(士)-氟比洛芬分子芳基基团的构象不利于其在流水座内结合,而(土)翻洛芬、(土)2苯基丙酸和(土)布洛芬分子芳基团的构象有利于其在疏水腔内结合。
底物结构的醇部分同样对反应速率有影响,M. Aroyo等1国在水饱和异辛统中采用固定化南极假丝酵母脂肪酶(Candida amartia)催化(土)-2-苯基丙酸与醉的酯化反应,发现伯醉反应速率最快,仲醇次之,叔醇几乎不反应J.M. Moreno等发现这主要是由于醇结合位点的宽度限制了仲醉和叔醇的结合.对于伯醇,短链的1-丙醇酯化速率要较1-辛醇和1-丁醇快。如果醇部分含有些拉电子基团, 如乙氧基、氨乙基、氰甲基等,由于其对酯键的活化,会明显提高水解反应的速率。如Bttistel等采用蔡普生乙氧基乙酯作为底物,Tai 等采用布洛芬氯乙基等,都明显提高了反应速率。
6:Candida rugosa 脂肪酶的同工酶
在许多微生物脂肪酶中,由于Candida rugosa脂肪酶具有高活性和立体选择性,因此对其研究也较深人。该脂肪酶在世界各地有许多生产商,过去一直认为他们的产品是均一的,但S. Allenmark等发现,Candida rugosa脂肪酶是由许多催化效率和立体选择性不同的同工酶组成的。在许多拆分反应中,这些同工酶的立体选择性都差别很大甚至相反,催化活性和稳定性差别也很大。R.C. Chang 等通过比较不同生产商提供的产品发现,同工酶各组分含量受培养条件、提纯方法等影响,由此导致了不同产品在催化效率和立体选择性上的差异。这大大限制了CRL在许多精细化学品合成中的应用,因此目前许多拆分反应都是采用生物化学纯化法或有机溶剂处理转化法获取的纯同工酶来进行的。




















.jpg)